
The authors of this content are Amy Charkowski, Kalpana Sharma, Monica L. Parker, Gary A. Secor, John Elphinstone
Potato Ring Rot Caused by Clavibacter michiganensis Subsp. sepedonicus
Taxonomy and Nomenclature
Clavibacter michiganensis subsp. sepedonicus is a Gram positive, coryneform, aerobic, non-spore-forming bacterium in the Microbacteriaceae family of the Actinobacteria. C. michiganensis is the only species currently recognized within the genus; all six of its subspecies (subspp. Insidiosus, michiganensis, nebraskensis, phaseoli, sepedonicus, and tesselarius) are plant pathogens.
C. michiganensis subsp. sepedonicus (Cms) was formerly known under the synonyms Corynebacterium sepedonicum, Corynebacterium michiganense pv. sepedonicum, and Corynebacterium michiganense subsp. sepedonicum.
Host Range
The only economically important host is potato (Solanum tuberosum), although natural infection was recently reported for the first time on tomato (van Vaerenbergh et al. 2016). Many members of the Solanaceae, including tomato and eggplant, are susceptible after artifical inoculation.
Some solanaceous weeds, e.g. hairy nightshade (Solanum sarrachoides) and buffalobur (S. rostratum), may harbor the bacterium following potato crops with ring rot (van der Wolf et al. 2005a).
The only economically important host is potato (Solanum tuberosum), although natural infection was recently reported for the first time on tomato (van Vaerenbergh et al. 2016). Many members of the Solanaceae, including tomato and eggplant, are susceptible after artifical inoculation.
Some solanaceous weeds, e.g. hairy nightshade (Solanum sarrachoides) and buffalobur (S. rostratum), may harbor the bacterium following potato crops with ring rot (van der Wolf et al. 2005a).
Geographical Distribution
First reported after an outbreak in Germany in 1905 (Appel 1906), it is one of the few major plant pathogens that is not present in the area where the crop evolved, i.e. Andean South America. In North America, it was first reported in Quebec (Canada) in 1931 and by 1940 it had spread to all important potato-producing districts in Canada and the USA due to movement in trade of infected seed potato tubers.
Subsequent zero tolerances imposed in quarantine and seed certification controls in most potato-growing areas have effectively limited the numbers of findings, although total eradication is difficult.
Currently, it tends to occur sporadically in cool, northern latitudes of North America (Northern USA and Canada) with only a single report in Mexico. Strict regulation in Europe has also reduced findings in annual surveys, especially in certified seed crops, with only occasional recent findings in some countries (Bulgaria, Czech Republic, Estonia, Finland, Germany, Greece, Hungary, Latvia, Lithuania, Netherlands, Norway, Slovakia, Sweden, and Turkey).
Isolated former outbreaks have been declared eradicated in Austria, Belgium, Cyprus, Denmark, France, Spain, and UK (England and Wales).
However, it remains prevalent in areas where formal seed certification is absent (parts of northern, western, and central Russia, Ukraine, Poland, and Romania). It is also reported in Asia (several provinces of China, Japan, Kazakhstan, Korea, Nepal, Pakistan, and Uzbekistan) although its distribution is not clearly defined.
Ring rot has never been confirmed in Africa, Australasia, or South America. In the field, foliar symptoms are not always observed or may occur only at the end of the season when they are difficult to distinguish in the senescing plant and are easily missed during crop inspections.
Unlike bacterial wilt, caused by Ralstonia, wilting due to the ring rot bacterium is usually slow and initially limited to the leaf margins (images below). Young infected leaves expand more slowly in the infected zones and become distorted.
Leaves affected by xylem blockages further down the stem often develop chlorotic, yellow to orange, interveinal areas. Infected leaflets, leaves, and even stems may eventually die. Leaves and tubers may simply be reduced in size and occasionally whole plants can be stunted.
First reported after an outbreak in Germany in 1905 (Appel 1906), it is one of the few major plant pathogens that is not present in the area where the crop evolved, i.e. Andean South America. In North America, it was first reported in Quebec (Canada) in 1931 and by 1940 it had spread to all important potato-producing districts in Canada and the USA due to movement in trade of infected seed potato tubers.
Subsequent zero tolerances imposed in quarantine and seed certification controls in most potato-growing areas have effectively limited the numbers of findings, although total eradication is difficult.
Currently, it tends to occur sporadically in cool, northern latitudes of North America (Northern USA and Canada) with only a single report in Mexico. Strict regulation in Europe has also reduced findings in annual surveys, especially in certified seed crops, with only occasional recent findings in some countries (Bulgaria, Czech Republic, Estonia, Finland, Germany, Greece, Hungary, Latvia, Lithuania, Netherlands, Norway, Slovakia, Sweden, and Turkey).
Isolated former outbreaks have been declared eradicated in Austria, Belgium, Cyprus, Denmark, France, Spain, and UK (England and Wales).
However, it remains prevalent in areas where formal seed certification is absent (parts of northern, western, and central Russia, Ukraine, Poland, and Romania). It is also reported in Asia (several provinces of China, Japan, Kazakhstan, Korea, Nepal, Pakistan, and Uzbekistan) although its distribution is not clearly defined.
Ring rot has never been confirmed in Africa, Australasia, or South America. In the field, foliar symptoms are not always observed or may occur only at the end of the season when they are difficult to distinguish in the senescing plant and are easily missed during crop inspections.
Unlike bacterial wilt, caused by Ralstonia, wilting due to the ring rot bacterium is usually slow and initially limited to the leaf margins (images below). Young infected leaves expand more slowly in the infected zones and become distorted.
Leaves affected by xylem blockages further down the stem often develop chlorotic, yellow to orange, interveinal areas. Infected leaflets, leaves, and even stems may eventually die. Leaves and tubers may simply be reduced in size and occasionally whole plants can be stunted.
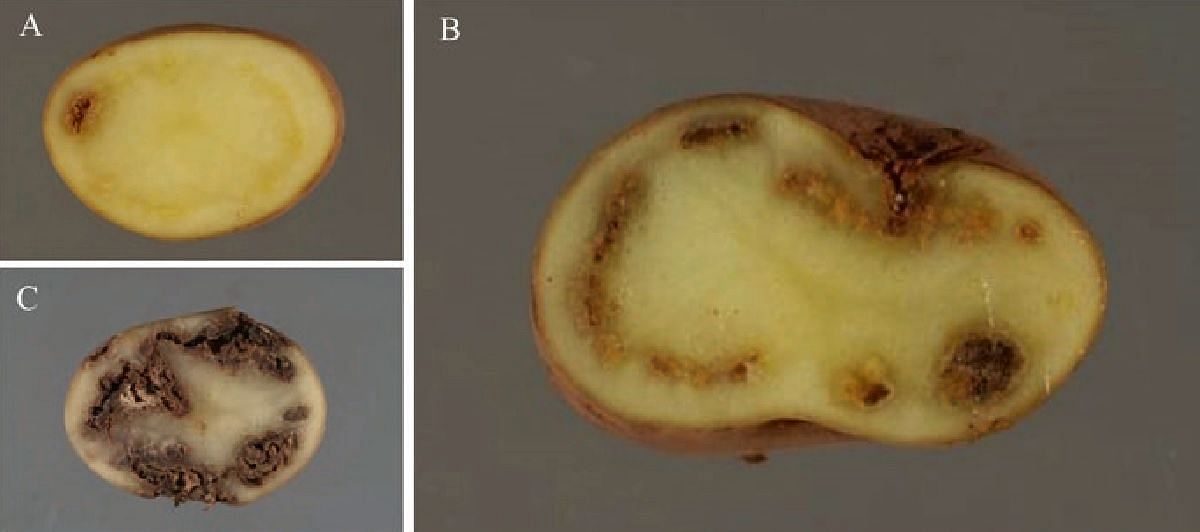
Symptoms of potato ring rot: initial tuber symptoms (water-soaked vascular ring, bacterial exudate, and start of vascular necrosis) (a), necrosis around vascular ring (b), advanced necrosis and secondary rotting (c). (Courtesy: Fera Science Ltd.)

Symptoms of potato ring rot: initial tuber symptoms interveinal chlorosis and wilting/epinasty at leaf margin (d, e), leaf distortion (f). (Courtesy: Fera Science Ltd.)
Epidemiology
Factors affecting the development and spread of potato ring rot were reviewed in detail by van der Wolf et al. (2005b). Seed potato tubers infected or contaminated with Cms are the primary source of infection. Inadvertent dissemination of the bacterium to new places of production occurs with the movement and planting of latently infected seed tubers.
The bacterium also spreads from infected tubers through direct contact and via contaminated surfaces of equipment used in potato production, such as seed cutters, planters, harvesters, graders, and transport vehicles as well as in contaminated stores and containers.
Plant-to-plant spread in the field is usually low but there is some experimental evidence that insects can transmit the disease (Christie et al. 1991) although the full significance of this is not understood. Cms survives for extended periods of many months to years in a dry and cool environment.
Its persistence on farm equipment, in stores, and on transport vehicles is an important means by which the bacterium is maintained and spread within farm units and disseminated to other production units. The bacteria migrate systemically from seed tubers to the stems via the vascular tissue, and subsequently into progeny tubers through the stolons.
The pathogen population density increases during the growing season but sometimes can be detected in stems within 3–4 weeks after planting infected seed. Survival of Cms in soil is not thought to contribute greatly to ring rot epidemiology although it can overwinter in the field in volunteer tubers (groundkeepers) and in potato tissue debris. Survival is longest in cold dry conditions.
The bacterium survives particularly well when dried in smears of decayed tuber tissue on equipment, machinery, potato sacks, and storage containers and can remain infectious in the dried state for at least 18 months at temperatures from 5 to −40 °C.
Cms has been reported to be associated with solanaceous weeds, but any role of these potential inoculum sources in the epidemiology of the ring rot disease of potato is unclear. Cms has a low optimum growth temperature (21–23 °C) and is confined mainly to cooler potato- growing regions.
Pathogenicity Determinants and Resistance
The genome of Cms was first sequenced by Bentley et al. (2008). In addition to the 3.26 Mb chromosome, which is highly similar amongst all Clavibacter subspecies, a 50-kb circular plasmid (pCS1) and a 90-kb linear plasmid (pCSL1) are carried by all Cms strains. The plasmid pCS1 is essential for symptom development, but genes required for host recognition, efficient colonization, infection, and evasion or suppression of plant defense are located on the chromosome (Eichenlaub and Gartemann 2011).
Two proteins, CelA and Chp-7, have been shown to be required for full virulence (Laine et al. 2000; Nissinen et al. 2001). The gene celA is located on the plasmid pCS1 and encodes the cellulase endo-β-1,4-glucanase. Located on the chromosome, chp-7 encodes a serine protease effector that directly elicits a hypersensitive response in nonhost tobacco plants (Lu et al. 2015).
Gene expression studies in Cms cells growing in either potato tissue or rich media (Holtsmark et al. 2008) have identified other putative virulence genes. In addition to celA, a homologous gene celB, two serine proteases and a xylanase was also upregulated in the plant tissue. Three other serine protease genes, including the chp-7, were downregulated.
Unlike Gram-negative bacterial pathogens of potato, the gram-positive Clavibacter pathogens do not have a type 3 secretion system (T3SS) to translocate effectors into host plant cells. Eichenlaub and Gartemann (2011) have described the likely infection process by Clavibacter. During infection, the bacteria enter the xylem vessels of host plants and subsequently spread systemically to colonize the whole plant.
Clavibacter can be considered as a biotrophic phytopathogen that recruits nutrients (carboxylic acids and sugars) from the xylem fluid. Following growth and colonization, the cell walls of the xylem vessels and surrounding parenchymatic cells are then hydolyzed by expression of cellulases and other extracellular enzymes, potentially including polygalacturonase, pectic lyase, xylanases, and other endoglucanases, leading to symptom development.
There are no currently available potato cultivars with immunity or useful resistance to ring rot. The concept of cultivar tolerance to ring rot is not yet understood and little is known of the status of most commonly grown cultivars with respect to their susceptibility to infection and colonization under varying environmental conditions.
Although potato cultivars vary in their propensities to express ring rot symptoms, less variation between cultivars in their susceptibilities to latent infections is observed (De Boer and McCann 1990). Since tolerant cultivars, which tend not to develop symptoms, can act as symptomless carriers of Cms, they have been removed from seed certification schemes in North America (Manzer and McKenzie 1988).
Laurila et al. (2003) demonstrated that an accession (PI472655) of the wild potato species Solanum acaule was susceptible to latent infection by Cms at 15 °C but appeared immune to infection at 25 °C.
The genome of Cms was first sequenced by Bentley et al. (2008). In addition to the 3.26 Mb chromosome, which is highly similar amongst all Clavibacter subspecies, a 50-kb circular plasmid (pCS1) and a 90-kb linear plasmid (pCSL1) are carried by all Cms strains. The plasmid pCS1 is essential for symptom development, but genes required for host recognition, efficient colonization, infection, and evasion or suppression of plant defense are located on the chromosome (Eichenlaub and Gartemann 2011).
Two proteins, CelA and Chp-7, have been shown to be required for full virulence (Laine et al. 2000; Nissinen et al. 2001). The gene celA is located on the plasmid pCS1 and encodes the cellulase endo-β-1,4-glucanase. Located on the chromosome, chp-7 encodes a serine protease effector that directly elicits a hypersensitive response in nonhost tobacco plants (Lu et al. 2015).
Gene expression studies in Cms cells growing in either potato tissue or rich media (Holtsmark et al. 2008) have identified other putative virulence genes. In addition to celA, a homologous gene celB, two serine proteases and a xylanase was also upregulated in the plant tissue. Three other serine protease genes, including the chp-7, were downregulated.
Unlike Gram-negative bacterial pathogens of potato, the gram-positive Clavibacter pathogens do not have a type 3 secretion system (T3SS) to translocate effectors into host plant cells. Eichenlaub and Gartemann (2011) have described the likely infection process by Clavibacter. During infection, the bacteria enter the xylem vessels of host plants and subsequently spread systemically to colonize the whole plant.
Clavibacter can be considered as a biotrophic phytopathogen that recruits nutrients (carboxylic acids and sugars) from the xylem fluid. Following growth and colonization, the cell walls of the xylem vessels and surrounding parenchymatic cells are then hydolyzed by expression of cellulases and other extracellular enzymes, potentially including polygalacturonase, pectic lyase, xylanases, and other endoglucanases, leading to symptom development.
There are no currently available potato cultivars with immunity or useful resistance to ring rot. The concept of cultivar tolerance to ring rot is not yet understood and little is known of the status of most commonly grown cultivars with respect to their susceptibility to infection and colonization under varying environmental conditions.
Although potato cultivars vary in their propensities to express ring rot symptoms, less variation between cultivars in their susceptibilities to latent infections is observed (De Boer and McCann 1990). Since tolerant cultivars, which tend not to develop symptoms, can act as symptomless carriers of Cms, they have been removed from seed certification schemes in North America (Manzer and McKenzie 1988).
Laurila et al. (2003) demonstrated that an accession (PI472655) of the wild potato species Solanum acaule was susceptible to latent infection by Cms at 15 °C but appeared immune to infection at 25 °C.
Significance and Economic Loss
Direct losses due to wilting and tuber rotting in field and store are usually moderate, especially where modern seed certification systems are in place. Nevertheless, ring rot constitutes a constraint on seed potato production, with considerable indirect effects on trade.
These result from statutory measures taken against ring rot outbreaks, which include loss of certification, restrictions on further cropping, purchase of new seed stocks, costs of disinfection, and disposal of infected and associated crops and subsequent effects on reputation and export trade.
Direct losses due to wilting and tuber rotting in field and store are usually moderate, especially where modern seed certification systems are in place. Nevertheless, ring rot constitutes a constraint on seed potato production, with considerable indirect effects on trade.
These result from statutory measures taken against ring rot outbreaks, which include loss of certification, restrictions on further cropping, purchase of new seed stocks, costs of disinfection, and disposal of infected and associated crops and subsequent effects on reputation and export trade.
Management
In the absence of effective chemical or biological control measures, or potato cultivars with adequate levels of resistance, management of potato ring rot must rely on the production and safe distribution of seed potatoes that are free from infection. Control is achieved primarily through strict application of quarantine and seed certification regulations, which involve a zero tolerance for the disease during seed and import inspections and for the pathogen during regular testing of consignments.
By laboratory testing for latent infections, infected lots can be detected early and eliminated from seed programs before the further spread of the pathogen occurs.
Phytosanitary measures must be aimed at the entire potato production system on account of the insidious nature of the disease. Seed potatoes should be imported only from countries which can show, by regular surveys and tests, that they operate a seed-potato production and distribution system free from ring rot.
Implementation of crop rotation, disinfection, and other sanitation practices is most important whenever the disease has occurred to prevent recurrence of the disease and spread of the pathogen. Ring rot infected crops and any adjacent crops that may have become contaminated should be eliminated from the production system and new certified seed should be acquired for any future production.
Disinfectants effective against Cms include quaternary ammonia-, chlorine-, or iodine-containing compounds. These should be applied, after cleaning of equipment and other contaminated surfaces, to ensure a minimum of 10 min contact under low organic load.
Control of potato volunteers and solanaceaous weeds is also important. The use of whole rather than cut seed helps to reduce any potential spread of the disease.
In the absence of effective chemical or biological control measures, or potato cultivars with adequate levels of resistance, management of potato ring rot must rely on the production and safe distribution of seed potatoes that are free from infection. Control is achieved primarily through strict application of quarantine and seed certification regulations, which involve a zero tolerance for the disease during seed and import inspections and for the pathogen during regular testing of consignments.
By laboratory testing for latent infections, infected lots can be detected early and eliminated from seed programs before the further spread of the pathogen occurs.
Phytosanitary measures must be aimed at the entire potato production system on account of the insidious nature of the disease. Seed potatoes should be imported only from countries which can show, by regular surveys and tests, that they operate a seed-potato production and distribution system free from ring rot.
Implementation of crop rotation, disinfection, and other sanitation practices is most important whenever the disease has occurred to prevent recurrence of the disease and spread of the pathogen. Ring rot infected crops and any adjacent crops that may have become contaminated should be eliminated from the production system and new certified seed should be acquired for any future production.
Disinfectants effective against Cms include quaternary ammonia-, chlorine-, or iodine-containing compounds. These should be applied, after cleaning of equipment and other contaminated surfaces, to ensure a minimum of 10 min contact under low organic load.
Control of potato volunteers and solanaceaous weeds is also important. The use of whole rather than cut seed helps to reduce any potential spread of the disease.